
In this image of marsh fox-tail grass (Alopecurus geniculatus) leaves, the arrow indicates the ligule at the junction of the blade and the sheath. The picture was taken by Christian Fischer, and is licensed under the Creative Commons Attribution-Share Alike 3.0 Unported license.
Having already started what might end up as a series of posts about underappreciated groups of plants [seaweed, and moss so far], I’ve decided to begin another about underappreciated plant parts. And I’m starting that collection with this item about a plant organ that’s very dear to my heart, the grass ligule. Or, more specifically, the membranous grass ligule.
Where our story begins…
For his MSc research at Durham University [located in the city of Durham, in the county of Durham, in the north-east of England, Michael Lee investigated the transport of radioactive amino acids along the leaf of meadow fescue (Festuca pratensis). Longish story short, he found that, in grass leaves that had been detached from the plant by cutting them where the leaf joined a node on the stem, the compounds travelled all the way from the cut base of the sheath towards the tip of the blade (Erin Garrett) in young leaves. But, in older leaves, upward transport of the amino acids was progressively restricted at the blade/sheath junction [B-SJ] [reminder, a grass leaf has two main components – the sheath, and the blade (Horace Leithead et al.)…].
Although one may ask questions about the relevance of this system to an intact grass plant, the observed ‘block’ at the B-SJ was deemed sufficiently intriguing to the Agriculture and Food Research Council [AFRC] [now part of the BBSRC [Biotechnology and Biological Sciences Research Council] for them to provide a substantial grant to study grass leaf senescence [a technical term for ageing (Matin Miryeganeh, Genes 2021, 12(2): 143; https://doi.org/10.3390/genes12020143). The task of undertaking that work fell to me as a Research Assistant at Durham University, working with the best PhD supervisor I could have wished for, Dr Alan Pearson. [For those who don’t know, in those days a Research Assistant was a member of staff who was paid to study for a PhD [Doctor of Philosophy award (Hasna Haidar; David Higginbotham)]. It really was the best of both words getting a salary whilst being a post-graduate ‘student’ and immersing oneself in a research topic for three years – happy times!].
However, and for reasons that aren’t essential to this story, we used Lolium temulentum (darnel) [for more on this fascinating grass, see Howard Thomas et al., Journal of Ethnobiology 36(1): 29-44, 2016; https://doi.org/10.2993/0278-0771-36.1.29; Howard Thomas, Plants People Planet 1: 197-203, 2019; https://doi.org/10.1002/ppp3.28%5D instead of F. pratensis. As for meadow fescue, darnel exhibited the same ‘amino acid blockage’ at the blade/sheath junction. That confirmation concentrated my interest in the anatomy of the cells, tissues, and organs at that region*, which, inevitably, led me to take a closer look at the ligule [which is where our story really begins].
What is the grass ligule?
Almost every source will define this structure as a membrane at the junction of the blade and the sheath of the grass leaf (Lizzie Harper; Jane Mangold; Daniel Murphy)**. Ligules of some species may have a variety of hairs or other structures associated with their outer surface or margins (Nigel Chaffey, Botanical Journal of the Linnean Society 89: 341–354, 1984; https://doi.org/10.1111/j.1095-8339.1984.tb02565.x). In some grasses the ligule may be reduced to a ring of hairs (Peter Landschoot); in others – e.g. Echinochloa crus-galli – it may be absent***. The variation in structure and appearance of their ligules has considerable value as an aid to the identification of grasses (Erin Garrett; Lizzie Harper; Jane Mangold; Fionnuala O’Neill) – particularly in the absence of flowers.
What does the ligule do?
The longest-held view of ligule function [attributed to Schlechtendahl by Ernst Hackel (page 4 in Die natürlichen Pflanzenfamilien. Teil II, Abteilung 2, Eds A Engler & K Prantl, 1887; Wilhelm Engelmann, Leipzig)] – the one you’ll usually find in textbooks and on the ‘net – is that it excludes water [memorably phrased as “a means to thwart water molecules from invading the sheath” (Nathan Claxton et al.)], dust and spores from entering the interior of the grass plant [e.g. Grahame Hubbard; JY Kim et al., Canadian Journal of Plant Science 97(6): 1068-1074, 2017; https://doi.org/10.1139/cjps-2017-0044%5D, and thereby stops the plant becoming water-logged or infected****. To my knowledge that proposal has never been tested experimentally. Rather, it’s a notion based upon the circumstantial evidence of the ligule’s position on the grass plant.
A deep dive into darnel ligule
The ligule of darnel is not much to look at with the naked eye or even with a hand-lens – it appears to be a whitish membrane-like structure situated at the junction between blade and sheath of a leaf, that’s a couple of millimetres tall [Fig. 1]. Because the sheath of the leaf to which it’s attached is closely pressed against the enclosed leaf or culm of the plant and partially wraps around it, the ligule – as an upward vertical extension of the sheath – also encircles the enclosed leaf or culm.

Fig. 1 When the blade of darnel is bent downwards the ligule is clearly seen as a wavy, translucent structure extending vertically upwards from the sheath. This is perennial ryegrass, which was used because I can’t find my pictures of darnel ligules. This image from Harry Rose is used under the Creative Commons Attribution 2.0 Generic license.
But, when prepared, sectioned, and examined under the light microscope [LM], it’s a revelation. In longitudinal section, the surface of the ligule that is next to the enclosed leaf/culm [and which is designated the adaxial [“referring to the side of an organ facing toward the axis”], which is the enclosed leaf/stem in this case] epidermis because it faces towards the stem of the plant] is continuous with the inner [adaxial] epidermis of the sheath. The outward-facing – abaxial – epidermis of the ligule is continuous with the adaxial epidermis of the blade. In transverse section, the ligule has a three-layered [tripartite] structure: both epidermides are uniseriate [i.e. a layer that is one cell thick], and a mesophyll [so-called because it’s the middle layer of a leaf-attached organ] that’s one to a few cells wide [widest towards the base of the ligule, thinnest towards the tip and margins]. Cells of the abaxial epidermis appear quite ‘empty’ indicating a large vacuole. Cells of the mesophyll and adaxial epidermis are progressively more heavily-stained indicating an increasing amount of cytoplasmic material/decreasing volume of vacuole within them [Fig. 2] (Nigel Chaffey, Annals of Botany 55: 65-75, 1985a; https://doi.org/10.1093/oxfordjournals.aob.a086879).
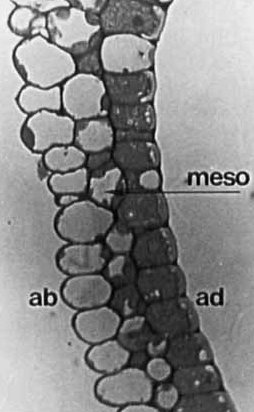
Fig. 2 Black-and-white light micrograph of Toluidine Blue-stained transverse section of membranous ligule of Lolium perenne [so long after the work was done I can’t find one of L. temulentum…]. The main feature to note is the marked increase in staining/reduction in proportion of unstained vacuole from abaxial epidermis (ab) to chloroplast-bearing mesophyll (meso), to the densely-cytoplasmic adaxial (ad) epidermis. Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x). Note: indications of scale and size of structures for this figure will be found in the publication from which the images are sourced. I apologise for not having included an appropriate scale bar for this figure in this post.
In the transmission electron microscope [TEM] (David Joy et al.), the view is even more impressive (Nigel Chaffey, Annals of Botany 55: 65-75, 1985a; https://doi.org/10.1093/oxfordjournals.aob.a086879) Not only was the degree of vacuolation observed with the LM confirmed, but – and as you should expect from this instrument – important information about type and number of organelles was also revealed. The highly-vacuolate cells of the abaxial epidermis had a correspondingly-thin cytoplasm layer that contained few organelles (and no noticeable chloroplasts), and a thick cuticle over the external cell walls. The moderately-vacuolated mesophyll cells are characterised by prominent populations of chloroplasts*****. The biggest surprise was the densely-cytoplasmic adaxial epidermal cells which had large numbers of mitochondria (Kara Rogers), and Golgi apparatus (or bodies) (Chris Hawes & Béatrice Satiat-Jeunemaitre, Trends in Plant Science 1: 395-401, 1996; https://doi.org/10.1016/S1360-1385(96)80315-3; Paul Dupree & D Janine Sherrier, Biochimica et Biophysica Acta 1404: 259-270, 1998; 10.1016/s0167-4889(98)00061-5), and abundant strands of rough endoplasmic reticulum [RER] [Fig. 3].

Fig. 3 TEM images of the adaxial epidermis of darnel ligule. Shown at left is the abundance of rough endoplasmic reticulum [RER]; on the right are several Golgi bodies and their empty-looking vesicles. Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x). Note: indications of scale and size of structures for this figure will be found in the publication from which the images are sourced. I apologise for not having included an appropriate scale bar for this figure in this post.
A cuticle was present over the outer cell walls of the adaxial epidermal cells, although this was thinner than that of the abaxial epidermis. Also of note was the abundance of plasmodesmata (Patricia Zambryski, Current Biology 18: PR324-R325, 2008; doi: https://doi.org/10.1016/j.cub.2008.01.046) in the walls between cells of the mesophyll and the adaxial epidermis. All of which catalogue of ultrastructure (see fig. 4) is not bad going for an organ that’s effectively written-off as a rather insignificant, small membranous structure.

Fig. 4 diagrammatic transverse section through the membranous ligule of darnel based on light and electron microscope observations. Abbreviations: abax, abaxial epidermis; adax, adaxial epidermis; ci, island of cytoplasm within cup-shaped depression of a plastid ; cl, chloroplast; cut, cuticle ; cv, coated vesicle ; d, dictyosome; dv, dictyosome-derived vesicle (not necessarily secretory); exp, extracellularly located product; mb, microbody; meso, mesophyll; mf, wall microfibril; ml, middle lamella; mt, mitochondrion; n, nucleus; nl, nucleolus; ol, oleosome; pd, plasmodesma; peri, periplasmic space; pl, plasmalemma; pmb, paramural body; po, polysome; pr, peripheral reticulum of chloroplast; pt, plastid ; rer, rough endoplasmic reticulum; ret, reticulate layer of cuticle ; st, starch; sv, secretory vesicle ; t, tonoplast; tr, transcellular strand; vac, vacuole; w, cell wall; 1, direct continuity between rough endoplasmic reticulum and cis aspect of dictyosome; 2, `smear’ of extracellularly located product over surface of cuticle ; 3, gaps in the cuticle. Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x).
As surprising as it [and it most certainly was!] was to find this degree of cellular complexity in the ligule, the puzzle now was to work out what it all meant. At first sight, it looks like this organ is photosynthetic, and has an energy-requiring synthesis of something in the abaxial layer which involves the Golgi and/or RER: But, what?
That ‘light-bulb moment’
And this is where serendipity [which I once saw defined as ‘looking for earthworms and finding gold’] comes into play. I’d read somewhere about use of optical brighteners****** (Dr Pete), specifically Calcofluor White to reveal cell walls in the fluorescence microscope (Toshiyuki Nagata & Itaru Takebe, Planta 92: 301–308, 1970; https://doi.org/10.1007/BF00385097; J Hughes & Margaret E McCully, Stain Technology 50: 319-329, 1975; doi: 10.3109/10520297509117082). This sounded like a nice and quick way to view cell arrangements and numbers of cell layers in thick-sections of ligules without all the fuss and time it took to produce semi-thin sections for light microscopy. It was. Using approx. 1 mm thick discs of material removed with a razor blade just above the B-SJ showed the ligule in transverse section and held in place between the base of its own leaf-blade and the enclosed leaf/stem. An example of the information this technique can give is shown in Fig. 5. Apart from the speed of preparation, a bonus of this procedure is that it avoids cellular collapse – especially of the highly-vacuolate adaxial epidermal cells – that often accompanies the aggressive fixation, etc. necessary for light and electron microscopy, i.e. the cell shapes are much more life-like (Nigel Chaffey, Botanical Journal of the Linnean Society 116: 53-69, 1994; https://doi.org/10.1111/j.1095-8339.1994.tb00421.x).
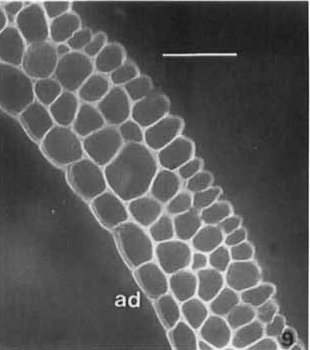
Fig. 5 Transverse section of ligule of Milium effusum with calcofluor, which nicely outlines the cells by ‘staining’ their walls [seen as white lines and curves], and shows their undeformed shapes. Note the relatively smooth outlines of the outer walls of the adaxial (ad) epidermal cells compared to the more ‘corrugated’ surface of the abaxial epidermis, which is likely to ensure a tighter seal of ligule to enclosed leaf/stem. Scale bar represents 50 μm. Image from Chaffey (Botanical Journal of the Linnean Society 116: 53-69, 1994; https://doi.org/10.1111/j.1095-8339.1994.tb00421.x).
The brightener worked with darnel ligules – when viewed in transverse section. However, occasionally bad cover-slip technique meant that a ligule came adrift and ended up flattened on the slide. Although that was no use for looking at number of cell layers, it showed an intriguingly-different fluorescence pattern to that seen in transverse sections. Basically, when looking at the adaxial surface of the ligule the fluorescence was confined to the area between the cell walls, i.e. the cells appeared as white fluorescing lozenges surrounded by a non-fluorescing boundary that corresponded to the position of the cell walls [Fig. 6].

Fig. 6 Adaxial surface of Lolium temulentum ligule with calcofluor. In this image the cell walls are ‘unstained’ and appear as dark boundaries to the cell contents that fluoresce white. Scale bar represents 50 μm. Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x).
What was causing that? We’ve had serendipity with the use of brighteners, now came the mini-eureka moment: The brightener appeared to be ‘staining’ the region between the outside of the cell membrane and the outer wall in the cells of the adaxial epidermis. Which area – the so-called periplasmic space (Vincent Franceschi & William Lucas, Protoplasma 107: 269–284, 1981; https://doi.org/10.1007/BF01276830; Tatiane Maria Rodrigues et al., Flora 206: 585-594, 2011; https://doi.org/10.1016/j.flora.2010.10.002) – was a region that had accumulations of circular profiles of membranes and other more fibrous looking material when seen in the TEM [the potential significance of which observation I hadn’t appreciated before hence its disclosure at this point in the story…]. Whatever the material was it appeared to have much greater affinity for the brightener than cellulose, but presumably had some chemical or physical characteristics in common with that cell wall polysaccharide to bind the calcofluor.
Could the material in the periplasmic space be something that the adaxial cells were synthesizing? If so, presumably it required energy – which might explain the large numbers of mitochondria in the cells. And, presumably also required the involvement of both the numerous Golgi bodies and abundant RER. Relating the carbohydrate-processing ability of the Golgi (Paul Dupree & D Janine Sherrier, Biochimica et Biophysica Acta 1404: 259-270, 1998; 10.1016/s0167-4889(98)00061-5), and the protein synthesis capacity of the RER, maybe the material was a glycoprotein – a molecule that has both a protein and a carbohydrate component?
A new function for the grass ligule?
Rather than the purely observational nature of the work to this point, we now had an hypothesis. Testing that hypothesis led to much more targeted work looking at the connection – both figuratively and quite literally – between the Golgi and the RER, the chemical nature of the accumulated product and a potential pathway of its synthesis suggested that what was being made was indeed a glycoprotein-like material synthesized in co-operation between Golgi and RER, and transported by vesicles to the periplasmic space [for more, see Nigel Chaffey, Protoplasma 127: 128-132, 1985b; https://doi.org/10.1007/BF01273709; Nigel Chaffey, Annals of Botany 76: 103-112, 1995; https://doi.org/10.1006/anbo.1995.1084%5D. But, the material didn’t appear to remain inside the ligule. In the TEM gaps in the cuticle of the adaxial epidermal cells could be seen with what looked like the material being released through the gaps to coat the outer surface of the cuticle [Fig. 7] – i.e. at the region between the ligule and the enclosed leaf/culm.

Fig. 7 Transmission electron micrograph of extracellular product in a cuticular gap and on the surface of the cuticle of an adaxial epidermal cell of darnel ligule. Image from Chaffey(New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x). Note: indications of scale and size of structures for this figure will be found in the publication from which the images are sourced. I apologise for not having included an appropriate scale bar for this figure in this post.
Since a secretory role for the ligule was rather at odds with the prevailing view of what it’s supposed to do, I was keen to get more support for this notion and chose to examine the root cap of darnel. Famously, this part of the plant produces so-called root-cap ‘slime’ (Robert Paull et al., Plant Physiology 56(2): 300–306, 1975; https://doi.org/10.1104/pp.56.2.300), or mucigel, or mucilage which is released from the cells and – amongst a number of roles proposed for this material (Toshihiro Watanabe et al., Plant Signal Behav. 3(8): 603-605, 2008; doi: 10.4161/psb.3.8.6356) – acts as a lubricant for the growth of roots through the soil. In many respects the cytochemistry, calcofluor-staining and assemblage of organelles found in the ligule’s adaxial epidermis was similar to that in the cells of the root cap of darnel (Nigel Chaffey, Annals of Botany 78: 3-13, 1996; https://doi.org/10.1006/anbo.1996.0088). Which was pretty good corroboration – and some sort of precedent – for my own view of ligule function.
Not one, but two functions for the ligule
After three years or so of reasonably intensive study, what did I conclude about membranous ligule function?*******
Two things mainly. First, the ligule of darnel could act passively to exclude water, dust and harmful spores – as has always been proposed. Whilst that was not novel, we now had an explanation of how this might work. When the highly-vacuolate abaxial epidermis cells are fully turgid [as they would be in life, and frequently fail to be after processing for the TEM] this helps the ligule to ‘curl’ around. In so doing, the outer walls of the adaxial epidermis are closely-appressed against the enclosed leaf/stem to give a pretty tight seal at that region.
But, it was apparent that the darnel ligule might also have a more active role. It looked like it was synthesising and secreting a glycoprotein-like substance to the outside of its adaxial epidermis [Fig. 8]. Because that is taking place at the blade/sheath region where the enclosing leaf is tightly pressed against the enclosed leaf, and by analogy with the root cap, it is proposed that the extracellular material might act as a lubricant that eases the exsertion of the enclosed leaf/culm.
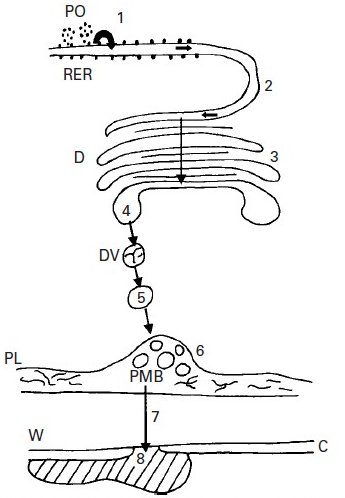
Fig. 8 Proposed route of synthesis, elaboration and secretion of the extracellular product of the adaxial epidermal cells of the ligule of Lolium temulentum. The protein component of the putative glycoprotein is synthesised on the ribosomes and polysomes (PO) and transferred to the lumen of the rough endoplasmic reticulum (RER) (1). Thence it is transported to the importing face of the Golgi [also known as dictyosome in plants ] (D) via the direct continuity between these two organelles (2). Further processing including the addition of carbohydrate to the RER-supplied product takes place during its passage across the Golgi (3). At the exporting face of the Golgi the ‘glycoprotein’ is packaged into smooth-membraned vesicles (DV) which bud off from the Golgi (4) and are transported to the outer cell membrane, the plasmalemma (PL) (5). At the plasmalemma the vesicles fuse to form paramural bodies (PMB) within the periplasmic space (6). The synthesised material is ultimately transported across the cell wall ![]() (7) and released to the outside of the ligule through gaps in the cuticle (C) over the outer tangential wall (8). Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x).
(7) and released to the outside of the ligule through gaps in the cuticle (C) over the outer tangential wall (8). Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x).
Is evidence from one species enough? Whilst it’s sufficient to challenge the orthodox view of a purely passive ligule function, it is always good to have supporting evidence from more than one taxon. Accordingly, I looked at a range of grasses (Nigel Chaffey, Botanical Journal of the Linnean Society 116: 53-69, 1994; https://doi.org/10.1111/j.1095-8339.1994.tb00421.x). Although I didn’t have time to study them as intensively as darnel, I had a look at their ultrastructure and behaviour with calcofluor. That study suggested that potentially secretory ligules are also found in seven other species: Bromus ramosus Huds., Festuca pratensis Huds., Lolium x hybridum Hausskn., L. multiflorum Lam., L. perenne L., Elytrigia repens L. Desv. ex Nevski ssp. repens [Fig. 9], and – possibly the most interesting of the species with such ligules, because its grain provides animal feed and supports 3 billion people worldwide – Triticum aestivum L., bread wheat.
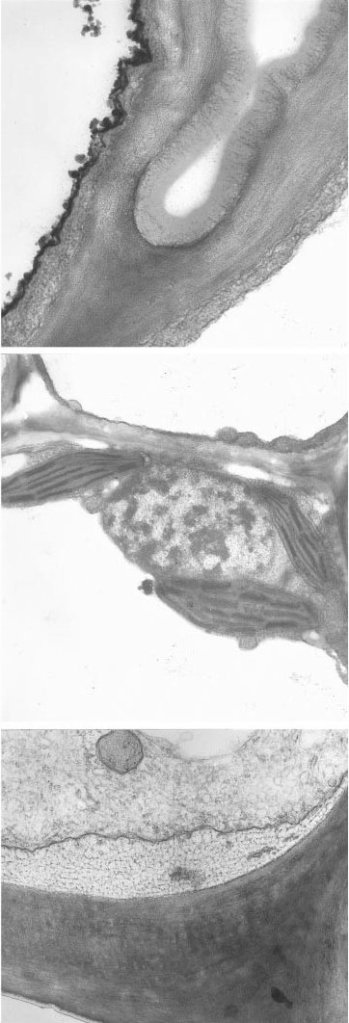
Fig. 9 Transmission electron micrographs of transverse section of the membranous ligule of Elytrigia repens ssp. repens illustrating its tripartite structure of, from top to bottom of sequence: empty-looking abaxial epidermis (with thick cuticle), chloroplast-bearing mesophyll, and the densely-cytoplasmic adaxial epidermis with fibrillar material in the periplasmic space. Image from Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x). Note: indications of scale and size of structures for this figure will be found in the publication from which the images are sourced. I apologise for not having included an appropriate scale bar for this figure in this post.
All good things come to an end…
Sadly, that was as far as I was able to get with my own studies of this fascinating plant organ. But, this was a great project to work on and gave me an in-depth knowledge of the minutiae of some grass ligules – and it was nice to find a little gold amongst the earthworms(!).
Well, anyway, that is why I find the grass ligule so fascinating, and why I’m always a little disappointed – although, sadly, not surprised – that nobody appears to have followed-up on this work. Certainly, ligule studies continue – e.g. rice (Dindin Hidayatul Mursyidin et al., Biodiversitas 22: 5280-5285, 2021; https://doi.org/10.13057/biodiv/d221207), Poaceae (Bruno Edson-Chaves et al., Bot. Rev. 89: 19–58, 2023; doi: http://dx.doi.org/10.1007/s12229-022-09285-3), and even in the Cyperaceae (Lucas Alves-dos-Santos et al., South African Journal of Botany 157: 372-379, 2023; https://doi.org/10.1016/j.sajb.2023.04.011) – but nobody seems to be looking at their function.
Oh well, that’s the way it goes. Thank you for reading this piece. And, if I’ve now made you look at grass ligules with a bit more admiration – and maybe even respect, they will be a little better-appreciated, and this post will have done its job.
* Although much of my time was spent looking at ligules, I never abandoned study of this ‘blockage’ and did find tyloses at the blade/sheath junction in older leaves of darnel [presence of which features were also hinted at by Lee’s MSc work on meadow fescue]. Tyloses are balloon-like extensions of parenchyma cells that poke into adjacent cells. In this instance they infiltrated the protoxylem lacunae [spaces in the vascular bundle that remain after the first-formed xylem vessels have been destroyed during development]. It was suggested that the tyloses might be removing the radioactive amino acids at this site which could explain the apparent block to further upward movement of these compounds during senescence of the darnel grass leaf [for more, see Nigel Chaffey & J Alan Pearson, Annals of Botany 56: 761-770, 1985; https://doi.org/10.1093/oxfordjournals.aob.a087068%5D.
** Other ligules exist, e.g. in sedges and rushes (Lizzie Harper), and other monocots (Paula Rudall & M Buzgo, 2002. Chapter 23 EvoIutionary history of the monocot leaf, pp. 431-458. In Developmental genetics and plant evolution (Eds QCB Cronk et al.), Taylor & Francis, London). The strap-shaped structure on the ray florets, or ray flowers, of composites (Lizzie Harper) is known as a ligule, as is a feature on the leaves of extant species of the lycophytes Selaginella and Isoetes. And ligules also refer to “bell-shaped structures within the girdle bands” of diatom frustules (Daniel Zuluaga-Astudillo et al., Appl. Sci. 2023, 13(1), 549; https://doi.org/10.3390/app13010549).
*** For reasons that I now forget, I also looked at the ligule of Agrostis gigantea, where I was surprised to find stomata-like structures in the abaxial epidermis (Nigel Chaffey, Annals of Botany 50: 717-720, 1982; https://doi.org/10.1093/oxfordjournals.aob.a086413). In the absence of verification that they were actually functional stomata (Regina Bailey), I erred on the side of caution and called them stomata-like, although they were only seen in association with vascular tissues in this ligule. Yes, you read correctly, some ligules have vascular tissue in them, which is continuous with the vascular tissue of the rest of the grass leaf. Which revelation means that some so-called membranous ligules are not simple ‘membranes’. I therefore use the term veined ligule to distinguish those ligules with vascular tissue – such as rice, marram, cock’s foot, Yorkshire fog, creeping bent, and black bent – from ligules that are entirely membranous – like those in darnel.
For more on veined ligules, see Nigel Chaffey, Annals of Botany 52: 13-21, 1983; https://doi.org/10.1093/oxfordjournals.aob.a086546; Nigel Chaffey, Botanical Journal of the Linnean Society 89: 341–354, 1984; https://doi.org/10.1111/j.1095-8339.1984.tb02565.x; Nigel Chaffey, New Phytologist 101: 613-621, 1985c; https://doi.org/10.1111/j.1469-8137.1985.tb02867.x – NB, the latter paper complicates matters by identifying heteroligulate species, grasses that bear both membranous and veined ligules.
**** Seemingly discounting that notion is Siva Chudalayandi who opines that a “more plausible explanation for ligules and auricles is that they might act as a pivot to help position the leaf blade at the correct angle to receive appropriate amounts of solar radiation”.
***** Discovery of the presence of chloroplasts in membranous ligules was a bit of a surprise. But, their presence raises the possibility that ligules may be photosynthetic organs. The narrow focus on ligule function proposed that mesophyll-produced photosynthetic products might fuel the energy-making activity of mitochondria for synthesis of the secretory product in the adaxial epidermal cells [remember the abundant plasmodesmata – channels that permit movement of material between cells – between the mesophyll and adaxial epidermis?]. However, the possibility that chloroplast-created sugars, etc. might be transported out of the ligule and may help the growth and development of the rest of the plant is also something to consider. To my knowledge that idea has never been followed-up – not even for human nutritionally-important wheat – despite evidence that photosynthesis of non-leaf plant parts can contribute to plant growth and development (Robert Henry et al., Biology 2020, 9(12): 438; doi: 10.3390/biology9120438).
But, other questions arise if such ligules are to photosynthesise. For example, how do the chloroplasts get CO2 through non-stomatous, thickly-cuticularised epidermis? In the absence of vascular connection with the rest of the leaf, how would any photosynthate get out from the ligule? And we don’t even know if the chloroplasts are photosynthetically active. They may look like photosynthetically-competent organelles, but are they functional? Proof that any piece of research should not only answer a question, but should raise others as well.
****** Readers of a certain age are probably familiar with optical brighteners because they were [still are..?] added to washing powders and detergents (Dr Pete) to give that brilliant blue-white appearance to washed clothing when held up against the daylight. This occurs because the brighteners bind to cellulose fibres – found in plant-derived materials such as cotton or linen – in the clothing and emit a bright whitish fluorescent light when exposed to ultra violet wavelengths. Which is why these compounds are particularly good at ‘staining’ the cellulose-rich cell walls of plants.
******* I could (should..?) have said right at the start of this article, if you’d like a summary of – most – of my thoughts on the membranous grass ligule, you could just look at Nigel Chaffey (New Phytologist 146: 5-21, 2000; https://doi.org/10.1046/j.1469-8137.2000.00618.x), which item is freely-accessible here, rather than wade through all of the other papers mentioned above. And, should anybody be that keen to know more, my PhD thesis is available here.

{kind=link}
{kind=link}
Leave a reply to Nigel Chaffey Cancel reply